
Théorie neutraliste en débat
dernière mise à jour le 19/12/2024
La théorie neutraliste de l’évolution moléculaire proposée par Motoo Kimura a mis l’accent sur l’importance des mutations neutres par rapport aux mutations adaptatives dans l’ADN. Cette théorie a dominé le paysage de l’évolution pendant un demi-siècle, mais elle continue de faire face à des défis.
Lorsque Charles Darwin a articulé sa théorie de l’évolution par sélection naturelle, il s’est concentré sur les adaptations – les changements qui permettent aux organismes de survivre dans des environnements nouveaux ou changeants. La sélection pour des adaptations favorables, a-t-il suggéré, a permis aux anciennes formes ancestrales de se diversifier progressivement en d’innombrables espèces.
Ce concept était si puissant que nous pourrions supposer que l’évolution est une question d’adaptation. Il peut donc être surprenant d’apprendre que pendant un demi-siècle, l’opinion dominante dans les cercles universitaires a été contraire à cette hypothèse.
Certes, la sélection ne fait aucun doute, mais de nombreux scientifiques ont fait valoir que la plupart des changements apparaissant au niveau du génome et sont essentiellement aléatoires et neutres. Les changements adaptatifs préparés par la sélection naturelle pourraient en effet transformer une nageoire en un pied primitif, mais ces changements ne contribuent que pour une faible part au processus évolutionniste où la composition de l’ADN varie le plus souvent sans aucune conséquence réelle.
Mais désormais, certains scientifiques s’opposent à cette théorie neutraliste, en disant que les génomes montrent beaucoup plus de preuves d’adaptation. Ce débat est important parce qu’il affecte notre compréhension des mécanismes qui génèrent la biodiversité, nos inférences sur la façon dont la taille des populations naturelles a changé au fil du temps et notre capacité à reconstruire l’histoire évolutive des espèces (y compris la nôtre). Ainsi, s’ouvre une nouvelle page de la science qui s’inspire du meilleur de la théorie neutraliste tout en reconnaissant l’influence réelle, empiriquement étayée de la sélection.
Une « fraction appréciable » de variation
L’idée centrale de Darwin était que les organismes présentant des caractéristiques désavantageuses seraient lentement éliminés par la sélection négative (ou purifiante), tandis que ceux ayant des caractéristiques avantageuses se reproduiraient plus souvent et transmettraient ces caractéristiques à la génération suivante (sélection positive). La sélection aiderait à diffuser et à affiner ces précieux traits. Pendant la majeure partie de la première moitié du XXe siècle, la génétique des populations s’est largement appuyée sur cette adaptation par sélection positive.
Mais en 1968, le célèbre généticien des populations Motoo Kimura a suggéré par sa théorie neutraliste qu’une « fraction appréciable » de la variation génétique au sein d’une espèce et entre espèces résulte d’une dérive génétique – c’est-à-dire des effets du hasard dans une population finie – plutôt que de la sélection naturelle, et que la plupart de ces différences n’ont pas de conséquences fonctionnelles sur la survie et la reproduction.
L’année suivante, Jack Lester King et Thomas Jukes ont publié un article célèbre qui soulignait également l’importance des changements génétiques aléatoires dans le cours de l’évolution. Un long débat a ensuite émergé entre neutralistes et adaptationnistes. Bien que tout le monde s’accorde à dire que la sélection purificatrice éliminerait les mutations délétères, les neutralistes étaient convaincus que la dérive génétique explique la plupart des différences entre les populations ou les espèces, tandis que les adaptationnistes les attribuaient à la sélection positive.
Une grande partie du débat a porté sur ce que Kimura entendait exactement par « fraction appréciable » Est-ce 50 %, 5 %, 0,5 %. On ne le savait pas, car Kimura parlait en qualitatif et non en quantitatif.
Néanmoins, la théorie neutraliste a été rapidement adoptée par de nombreux biologistes. En partie par la réputation de Kimura reconnu comme l’un des généticiens des populations théoriques les plus éminents de l’époque, mais également par des données mathématiques assez simples et intuitives.
Pour appliquer un modèle neutre d’évolution à une population, il n’est pas nécessaire de savoir à quel point la sélection est forte, quelle est la taille de la population, si les mutations sont dominantes ou récessives, ou si les mutations interagissent avec d’autres mutations. Dans cette théorie neutraliste, tous ces paramètres, très difficiles à estimer, disparaissent.
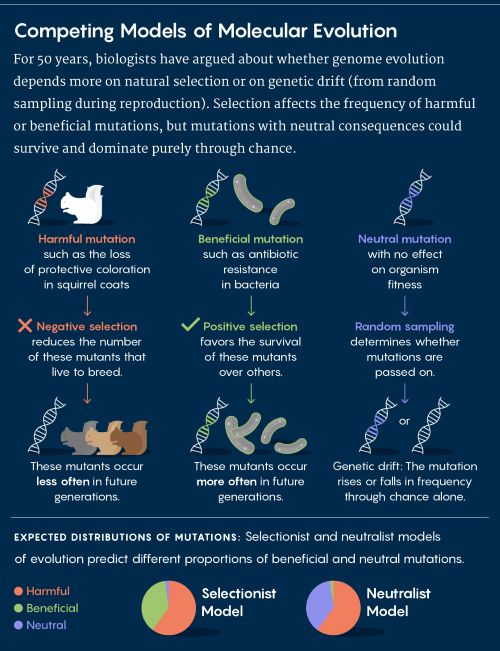
Figure 1 :
Pendant un demi-siècle, les biologistes de l’évolution ont débattu pour savoir si l’évolution du génome dépend davantage de la sélection naturelle ou de la dérive génétique (issue d'un processus aléatoire au cours de la reproduction)
La sélection affecte la fréquence des mutations néfastes ou bénéfiques, mais les mutations aux conséquences neutres peuvent survivre et dominer uniquement par le hasard.
La seule clé d’entrée requise par le modèle neutraliste est le produit de la taille de la population par le taux de mutation par génération. À partir de ces informations, le modèle neutre peut prédire comment la fréquence des mutations dans la population changera au fil du temps. En raison de sa simplicité, de nombreux chercheurs ont adopté ce modèle comme une explication par défaut des modèles de variation génétique qu’ils avaient observés.
Cependant, maints généticiens des populations n’ont pas été convaincus, car certains modèles basés sur la sélection naturelle pouvaient expliquer les modèles observés dans la nature, aussi bien que le modèle neutraliste, si ce n’est mieux.
Plus fondamentalement, même lorsqu’il n’y a pas assez de données pour réfuter le modèle neutre, cela ne signifie pas l’absence de la sélection naturelle.
Pendant des décennies, cela a été le nœud du problème : Kimura avait proposé une théorie neutraliste à une époque où les technologies de séquençage et la réaction en chaîne par polymérase n’étaient pas disponibles. Nous n’en savions pas assez sur la variation génomique pour résoudre le différend.
Des sentiments forts à l’égard de la neutralité
Aujourd’hui, 50 ans après l’article de Kimura, un séquençage génomique plus abordable et des méthodes statistiques sophistiquées permettent aux théoriciens de l’évolution de mieux quantifier chaque contribution, neutre ou adaptative, à l’évolution des populations et des espèces. Chez des espèces comme les humains et les mouches des fruits, les données ont révélé une sélection et une adaptation étendues, ce qui a conduit à une forte résistance à l’idée originale de Kimura, du moins par certains chercheurs.
Bien que la grande majorité des chercheurs s’accordent à dire que la stricte neutralité telle qu’elle a été formulée à l’origine est fausse, beaucoup soulignent également que les raffinements de la théorie neutraliste ont permis de remédier à ses faiblesses. L’une des lacunes initiales était qu’elle ne pouvait pas expliquer les différents modèles d’évolution du génome observés entre des espèces aux tailles de population différentes. Par exemple, les espèces dont la population est plus petite ont en moyenne plus de mutations délétères.
Pour y remédier, Tomoko Ohta, l’une des élèves de Kimura, a proposé une théorie suggérant que de nombreuses mutations ne sont pas strictement neutres, mais légèrement délétères. Elle a fait valoir que si la taille des populations est suffisamment grande, la sélection purificatrice les purgera des mutations, même légèrement délétères. Dans les petites populations, cependant, la sélection purificatrice est moins efficace et permet aux mutations légèrement délétères de se comporter de manière neutre.
Cette théorie « presque neutraliste » posait cependant problèmes. Elle n’expliquait pas, par exemple, pourquoi le taux d’évolution varie comme observé entre différentes lignées d’organismes. Face à ces défis, Ohta a développé une autre variante du modèle presque neutre en 1990. Malgré tout, cette nouvelle théorie ne fait guère progresser le débat.
Dans quelle mesure on évolue de manière neutre ?
Le débat en cours entre neutralistes et sélectionnistes n’est pas particulièrement fructueux. On en revient à la question quantitative de la part de la sélection. En réalité, certains sites sont complètement neutres, certains sites sont modérément sélectionnés, et d’autres sont fortement sélectionnés.
Lorsque Townsend a commencé à étudier le cancer il y a une dizaine d’années après une formation de biologiste de l’évolution, il a constaté que les biologistes du cancer avaient commencé à étudier les mutations à un niveau de détail qui pouvait révéler des informations sur les taux de mutation à des sites individuels du génome. C’est une information précieuse que la plupart des généticiens des populations n’obtiennent pas des populations sauvages qu’ils étudient. Pourtant, peu de biologistes du cancer étudient la sélection naturelle, et c’est ce que Townsend a apporté au domaine du cancer avec sa formation en biologie évolutive.
Il a publié les résultats d’une analyse évolutionniste des mutations dans les cancers où il quantifie site par site, quelles sont les intensités de sélection des différentes mutations. Les cellules cancéreuses regorgent de mutations, mais seul un petit sous-ensemble d’entre elles est fonctionnellement important pour le cancer. Les intensités de sélection révèlent l’importance des différentes mutations pour stimuler la croissance dans un cas individuel de cancer – et donc celles qui seraient les plus prometteuses en tant que cibles thérapeutiques.
Bien que l’identification des mutations subissant la sélection la plus forte soit clairement utile et importante, la sélection peut également avoir des effets indirects subtils mais importants sur les régions du génome voisines de la cible de la sélection.
Le premier indice de ces effets indirects est apparu dans les années 1980 et 1990 avec l’avènement de la réaction en chaîne par polymérase, une technique qui a permis aux chercheurs d’examiner pour la première fois la variation au niveau des nucléotides dans les séquences génétiques. Ils ont découvert une corrélation apparente entre le niveau de variation génétique et le taux de recombinaison à n’importe quelle région spécifiée du génome.
La recombinaison est un processus dans lequel les copies maternelles et paternelles des chromosomes échangent des blocs d’ADN entre elles pendant la méiose, la production de spermatozoïdes et d’ovules. Ces recombinaisons mélangent la variation génétique dans tout le génome, divisant des allèles qui auraient pu être auparavant ensemble.
En 2005, les chercheurs ont pu obtenir des données sur l’ensemble du génome d’une variété d’organismes, et ils ont commencé à trouver partout cette corrélation apparente entre les niveaux de variation génétique et les taux de recombinaison. Cette corrélation signifiait que des forces allant au-delà de la sélection de purification directe et de la dérive neutre créaient des différences dans les niveaux de variation dans le paysage génomique.
Les différences dans les taux de recombinaison à travers le génome révèlent un phénomène appelé auto-stop génétique. Lorsque les allèles bénéfiques sont étroitement liés à des mutations neutres voisines, la sélection naturelle a tendance à traiter ces parties liées comme une unité.
L’auto-stop génétique oblige les généticiens évolutionnistes à une toute nouvelle force évolutionniste, celle de la « sélection liée ». Si seulement 10 % du génome est soumis à la sélection directe dans une population, alors la sélection liée signifie qu’un pourcentage beaucoup plus important – peut-être 30 ou 40 % – peut montrer ses effets.
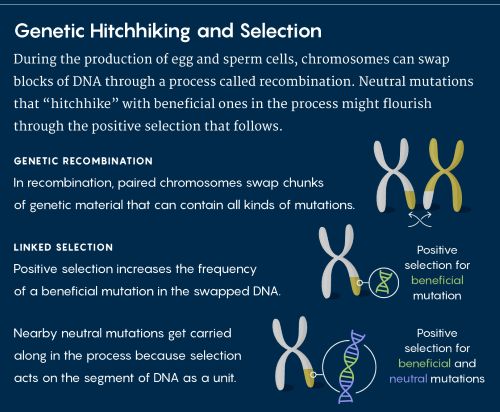
Figure 2 :
Pendant la recombinaison des chromosomes au moment de la fécondation, les mutations neutres prises en « autostop » par des mutations bénéfiques pourraient prospérer par la sélection positive qui s’ensuit.
Et si cela est vrai, alors la sélection des variants adaptatifs façonne indirectement les régions génomiques voisines, conduisant à ce que la fréquence des allèles ne soit pas seulement liée à la dérive génétique, mais aussi à une nouvelle couche de stochasticité induite par la sélection. La sélection liée produirait plus de variance entre les générations que ce à quoi on pourrait s’attendre avec la neutralité.
Les mutations neutres liées restent neutres. Elles pourraient faire de l’auto-stop avec des allèles bénéfiques, mais cette liaison est aléatoire. Elles pourraient tout aussi bien être liés à des allèles nuisibles et éliminés par une « sélection de fond ». Ainsi, le destin des mutations neutres est toujours déterminé par le hasard.
Certes, les mutations neutres sont toujours neutres – mais elles ne se comportent pas comme la théorie neutraliste le prédirait. La sélection purificatrice sur des sites liés va au-delà de la simple dérive, tandis que la sélection de fond et l’auto-stop conduisent à moins de variation génétique que sous neutralité.
Modèles neutres et évolution humaine
Si les modèles neutres ont sans aucun doute produit d’énormes fruits théoriques, le pouvoir explicatif de la théorie neutraliste n’a jamais été exceptionnel. Cinq décennies après, à l’ère du séquençage facile et des énormes ensembles de données génomiques de population, le pouvoir explicatif de la théorie neutraliste semble encore pire.
Chez l’homme, des preuves récentes suggèrent qu’il y a beaucoup plus d’adaptation que nous ne le pensions. L’évolution humaine récente est en grande partie une histoire de migrations vers de nouveaux lieux géographiques où les humains ont rencontré de nouveaux climats et agents pathogènes auxquels ils ont dû s’adapter. En 2017, Kern a publié une étude montrant que la plupart des adaptations humaines résultent d’une variation génétique existante dans le génome, et non de nouvelles mutations qui se propagent rapidement dans la population.
Même si seulement environ 1% du génome humain code réellement pour les protéines, peut-être qu’environ 20 % du génome régule quand et où ces régions codantes sont exprimées. Mais cela laisse encore environ 80% du génome avec une fonction inconnue.
Certaines de ces parties non codantes du génome sont criblées de séquences d’ADN répétitives, causées par des éléments génétiques transposables, ou transposons, qui se copient et s’insèrent dans tout le génome. Cette partie du génome est essentiellement neutre au sens de Kimura, même si une fraction de ces transposons affecte l’expression des gènes. Pour cette raison, certains pensent que des modèles neutres appliqués aux régions non fonctionnelles du génome peuvent être utilisés pour déduire l’histoire démographique des populations humaines avec une grande précision. D’autres, inversement, pensent qu’en y introduisant la sélection, ces modèles de simulation démographique échouent.
Kern pense que la sélection liée est importante, mais nous ignorons toujours quelle est l’ampleur de sa fraction dans le génome. Un article récent estime que cette sélection de fond est portée par une fraction qui peut concerner jusqu’à 80 à 85% du génome humain.
Après avoir également pris en compte les changements biaisés dans les gènes que la recombinaison peut introduire lors de la réparation de l’ADN, ils ont conclu que moins de 5% du génome humain a évolué par le seul hasard. Cela suggère que si la plupart de notre matériel génétique est formé de séquences non fonctionnelles, la grande majorité de celui-ci évolue indirectement sous l’effet d’un certain type de sélection.
Il est possible que cette estimation augmente à mesure que les biologistes apprendront à reconnaître des indices plus subtils de sélection. La nouvelle frontière de la génomique des populations se concentre sur des traits tels que la taille, la couleur de la peau et la pression artérielle (parmi beaucoup d’autres) qui sont polygéniques, ce qui signifie qu’ils résultent de centaines ou de milliers de gènes agissant de concert. La sélection pour une plus grande taille, par exemple, nécessite d’empiler des changements sur un certain nombre de gènes dispersés pour avoir un effet. De même, lorsque les agriculteurs sélectionnent des souches de maïs pour des rendements plus élevés, l’impact se manifeste généralement dans de nombreux gènes simultanément.
Mais la détection des adaptations polygéniques dans les populations naturelles est une « affaire très délicate », car ces multitudes de gènes sont susceptibles d’interagir de manière complexe et non linéaire. Les méthodes statistiques permettant de repérer ces séries de changements commencent à peine à être développées. Il nous faudra donc apprendre à reconnaître de « nouvelles saveurs » de l’adaptation, comme de nombreux petits changements dans les fréquences de mutation individuelles qui comptent collectivement pour la sélection naturelle.
En d’autres termes, il s’agit d’un autre mécanisme non neutre affectant l’évolution du génome. Aussi utile que la théorie neutraliste ait été sous ses diverses formes au cours du dernier demi-siècle, l’avenir des sciences de l’évolution dépendra de la découverte des processus toujours plus raffinés par lesquels la sélection façonne inexorablement nos génomes.
Bibliographie
Callier V
Theorists Debate How ‘Neutral’ Evolution Really Is
https www quantamagazine org neutral theory of evolution challenged by evidence for dna selection 20181108
Cannataro VL, Gaffney SG, Townsend JP
Effect Sizes of Somatic Mutations in Cancer
J Natl Cancer Inst. 2018 Nov 1;110(11):1171-1177
DOI : 10.1093/jnci/djy168
Kimura M
Evolutionary rate at the molecular level
Nature. 1968 Feb 17;217(5129):624-6
DOI : 10.1038/217624a0
King JL, Jukes TH
Non-Darwinian evolution
Science. 1969 May 16;164(3881):788-98
DOI : 10.1126/science.164.3881.788
Ohta T
Slightly deleterious mutant substitutions in evolution
Nature. 1973 Nov 9;246(5428):96-8
DOI : 10.1038/246096a0
Pouyet F, Aeschbacher S, Thiéry A, Excoffier L
Background selection and biased gene conversion affect more than 95% of the human genome and bias demographic inferences
Elife. 2018 Aug 23;7:e36317
DOI : 10.7554/eLife.36317
Schrider DR, Kern AD
Soft Sweeps Are the Dominant Mode of Adaptation in the Human Genome
Mol Biol Evol. 2017 Aug 1;34(8):1863-1877
DOI : 10.1093/molbev/msx154
Zhang J
Neutral Theory and Phenotypic Evolution
Mol Biol Evol. 2018 Jun 1;35(6):1327-1331
DOI : 10.1093/molbev/msy065
Et pour aller plus loin
Profil de nos 5000 abonnés
| Par catégorie professionnelle | |
| Médecins | 27% |
| Professions de santé | 33% |
| Sciences de la vie et de la terre | 8% |
| Sciences humaines et sociales | 12% |
| Autres sciences et techniques | 4% |
| Administration, services et tertiaires | 11% |
| Economie, commerce, industrie | 1% |
| Médias et communication | 3% |
| Art et artisanat | 1% |
| Par tranches d'âge | |
| Plus de 70 ans | 14% |
| de 50 à 70 ans | 53% |
| de 30 à 50 ans | 29% |
| moins de 30 ans | 4% |
| Par motivation | |
| Patients | 5% |
| Proche ou association de patients | 3% |
| Thèse ou études en cours | 4% |
| Intérêt professionnel | 65% |
| Simple curiosité | 23% |
Si vous n'êtes pas abonné
C'est ici
INUTILE si vous vous êtes déjà abonné, car vous le restez tant que vous ne demandez pas votre désisncription
Médecine évolutionniste (ou darwinienne)
Depuis quelques années, le problème de l'antibiorésistance, les progrès de la génomique, la redécouverte du microbiote et la prise en charge de maladies au long cours, nécessitent l'introduction d'une pensée évolutionniste dans la réflexion clinique.
Le premier diplôme universitaire intitulé "Biologie de l'évolution et médecine" a été mis en place à la faculté de Lyon en 2016.
Vous aimerez aussi...
ADN poubelle de l'intelligence humaine - Les traits qui séparent les chimpanzés des humains semblent évidents, nous sommes pourtant des [...]
Latéralisation droite de la main chez l'homme. - Préhistoire de la latéralisation droite : archéologie et éthologie. [...]
Un virus géant qui a façonné l'arbre de vie - Cette étude a identifié une famille de virus géants qui a peut-être joué un rôle clé dans [...]
Césariennes, flore intestinale et obésité. - Médecins et épidémiologistes avaient constaté depuis longtemps que la naissance par césarienne [...]
Gène de fécondité hérité de Néandertal - Le récepteur de la progestérone de Néandertal a un effet favorable sur la [...]
Vous aimerez aussi ces humeurs...
Pailles de la prévention - Les jours de grande pollution, de nombreux messages officiels recommandent de garder les jeunes [...]
La cigarette au cinéma - Les producteurs de films ont une obsession budgétaire bien compréhensible. La [...]
Manipulations grossières du ministère de la santé - Une publicité en faveur du dépistage de masse, entendue sur les médias grand public, [...]
Deux histoires - La première histoire se passe en mars 1970. Un jeune médecin fraichement installé dans un [...]
Adage d'hiver - « Un rhume dure une semaine sans soins et 7 jours avec soins ». Les adages sont le vif [...]
Vous aimerez aussi...
Microbiote du syndrome du côlon irritable - Les bactéries intestinales altèrent le fonctionnement de l'intestin et du [...]
Notre cœur est adapté à des efforts constants - Contrairement à d’autres grands singes, les humains ont développé des capacités [...]
Influence de l’histoire évolutionniste sur la santé et les maladies - En glissant avec la souris sur les mots surlignés, leur explication apparaît. Abstract Presque [...]
Les religions ont augmenté la taille des sociétés - Il y a beaucoup de choses qui permettent aux humains de vivre en grands groupes et de s'entasser [...]
Différences sexuelles du cerveau des rongeurs - Abstract Les hormones stéroïdes d'origine gonadique agissent sur le cerveau néonatal pour [...]