
Écologie évolutionniste de la vie humaine
dernière mise à jour le 26/04/2025
Abstract
L’histoire de la vie humaine possède plusieurs caractéristiques inhabituelles, notamment les gros bébés, la puberté et la ménopause tardives, et le fait qu’il existe une forte influence culturelle sur les décisions reproductives tout au long de la vie. Dans cette revue, j’examine l’histoire de la vie humaine d’un point de vue écologique évolutionniste. J’examine d’abord les preuves des compromis de l’histoire de vie entre la fertilité et la mortalité chez l’homme. Les modèles de croissance, de fécondité et de mortalité tout au long de la vie sont ensuite discutés et illustrés par des données provenant d’une population gambienne traditionnelle. Après avoir décrit les étapes du parcours de la vie humaine, je discute de deux phénomènes d’un intérêt particulier en anthropologie évolutionniste, qui sont tous deux apparemment uniques aux humains et qui ne sont pas encore entièrement compris. Tout d’abord, je discute de l’évolution de la ménopause, donc de l’arrêt de la reproduction féminine bien avant la mort. J’examinerai les preuves de cette évolution par l’hypothèse que l’investissement dans le succès reproductif futur des enfants existants est plus important que la poursuite de la procréation jusqu’à un âge avancé sont examinées, ainsi que les données relatives aux contraintes biologiques qui peuvent intervenir. Deuxièmement, je discute de la transition démographique. La baisse de la fécondité à une époque où les ressources sont de plus en plus abondantes représente un sérieux défi pour une vision évolutionniste de l’histoire de la vie et du comportement humains, cela sera donc examinée en détail. L’investissement des parents dans la compétition entre les enfants peut être la clé pour comprendre ces deux phénomènes humains inhabituels.
Les humains ont des bébés gros, mais relativement vulnérables
La mortalité infantile est élevée, la rivalité entre frères et sœurs est féroce, la puberté est tardive, et, pour les femmes, les grossesses sont rapides et dangereuses, les efforts parentaux sont pénibles et longs, et la vie post-reproductive est longue. Si nombre de ces caractéristiques sont communes aux grands singes, beaucoup sont uniques à Homo sapiens ou particulièrement exagérées.
Les humains sont si inhabituels à bien des égards qu'il est difficile d'attribuer un aspect de notre comportement ou de notre histoire biologique à un aspect de notre écologie par rapport aux autres primates. Une particularité remarquable est la taille du cerveau, qui se situe deux à trois écarts types au-dessus de la ligne prédisant la taille du cerveau par rapport à la taille du corps chez les primates[1] ; et il est plausible (bien que loin d'être certain) que de nombreuses autres caractéristiques inhabituelles de l'histoire biologique humaine y soient liées. Les conséquences d'un cerveau volumineux sont à la fois comportementales et physiologiques.
Dans cette analyse, je me concentre principalement sur les aspects de l'histoire de la vie humaine qui sont comportementaux et qui sont le plus liés à notre système social. Or, notre système social a exercé une influence considérable sur l'évolution de notre physiologie et de notre psychologie.
Le comportement humain est fortement influencé par la culture, elle-même un sous-produit de notre intelligence. Comportement et culture sont si étroitement liés que j'utilise ces deux termes presque comme synonymes dans cette analyse. Nous pouvons décider quand nous accoupler et avoir des enfants, quelle quantité de nourriture leur donner et combien de temps vivre avec eux. Il s'agit soit de décisions individuelles, soit de comportements adoptés par souci de respecter une norme culturelle ; la plupart de ces décisions sont une combinaison de ces deux caractéristiques, qui sont sans doute également influencées par la génétique.
Parmi ceux qui étudient l'évolution du comportement humain, les points de vue divergent quant au cadre théorique le plus susceptible de nous aider à comprendre nos actions. Les psychologues évolutionnistes estiment qu'une grande partie de notre comportement peut être interprétée comme une adaptation génétique à un environnement ancestral dans lequel nous avons vécu en tant que chasseurs-cueilleurs pendant la majeure partie de notre histoire évolutive. Cet environnement ancestral est souvent qualifié d'environnement d'adaptation évolutionniste, généralement présenté comme une savane africaine sèche. Ce modèle s'inspire largement de l'un des premiers groupes de chasseurs-cueilleurs groupes dont l'histoire biologique a été étudiée : les !Kung d'Afrique australe[2]. Ce peuple doux, monogame et musicien, aux naissances soigneusement espacées, constitue un candidat intéressant pour l'unité sociale ancestrale humaine typique. Mais les légendes sur la vie des !Kung s'estompent. Leur célèbre intervalle de naissance de 4 ans est passé à 2,5 à 3 ans, plus courant, lorsque les !Kung étaient traités pour des maladies sexuellement transmissibles[3] comme c'était le cas pour toutes les autres populations du nord-ouest du Botswana. Et dans l'étude de l'histoire de la population humaine menée par Cavalli-Sforza en 1994, les !Kung sont décrits comme un candidat improbable pour un groupe africain ancestral, qui pourrait même avoir migré d'Asie vers l'Afrique. Des études ultérieures sur d'autres chasseurs-cueilleurs ont révélé ce que tout écologiste comportemental aurait pu soupçonner : l'absence d'écologie humaine typique. Une étude récente sur les Ache du Paraguay[4], les auteurs décrivent un groupe qui, à bien des égards, est bien plus susceptible de caractériser l'humanité ancestrale. Les Ache ont été beaucoup moins exposés aux groupes sédentaires que les !Kung. Habitants de la forêt, ils ont vécu dans un isolement extrême jusqu'en 1972, année où ils ont été dévastés par des épidémies transmises par les missionnaires. Ceux qui ont survécu ont dû se retirer dans des réserves. Leur histoire a révélé une vie de difficultés, de violence au sein des groupes sociaux et entre eux, et une reproduction rapide. L'infanticide et le géronticide étaient courants, les groupes familiaux instables et peu d'individus vivaient assez longtemps pour contribuer de manière significative à l'unité sociale après la reproduction. Des guerres constantes entre groupes, avec des populations parfois presque anéanties, parfois en expansion rapide vers de nouvelles zones, pourraient bien être les circonstances dans lesquelles notre cycle biologique a évolué.
Certains psychologues évolutionnistes[5] ont soutenu que l'origine de l'agriculture, qui a alimenté l'expansion démographique, est si récente que l'évolution n'a pas eu le temps de modifier nos comportements depuis. Il est donc peu utile d'étudier les conséquences sur la fitness de tout comportement dans des sociétés autres que celles des chasseurs-cueilleurs. Cependant, il existe des preuves génétiques d’adaptation chez l'homme depuis l'origine de l'agriculture ; par exemple, la capacité à digérer le lactose à l'âge adulte a évolué en tant qu'adaptation à la production laitière chez des groupes qui élèvent du bétail depuis moins de 10 000 ans[6],[7]. On peut affirmer de manière convaincante que les charges de travail, l'expérience reproductive et bien d'autres aspects du cycle biologique n'ont pas autant changé lors de la transition entre la cueillette et l'agriculture paysanne qu'ils ne l'ont fait beaucoup plus récemment avec la transition démographique vers des familles de petite taille[8], un sujet sur lequel je reviens plus loin.
Étant donné que nous ignorons ce que faisaient nos ancêtres, nous ne pouvons pas nous comparer de manière significative aux autres primates, et comme les humains sont généralement peu sensibles aux manipulations expérimentales, il nous reste à examiner les corrélats écologiques et sociaux des variations individuelles et interpopulationnelles du cycle biologique humain.
Les méthodes d'écologie comportementale et évolutionniste sont donc probablement importantes. Pour les êtres humains, l'environnement culturel est une donnée aussi importante que toute autre. La culture fait partie intégrante de notre environnement. Les outils de l'écologie comportementale doivent donc être appliqués dans un cadre où l'environnement inclut la culture. Certains anthropologues évolutionnistes ont étudié l'évolution culturelle en modélisant la transmission d'unités culturelles, ou « mèmes », comme des unités héréditaires distinctes soumises à la sélection, bien que présentant des caractéristiques de transmission génétiques différentes[9]. Tout en reconnaissant que la culture est assez différente des autres aspects de l'environnement, cette approche n'a pas offert beaucoup d'opportunités pour tester empiriquement des hypothèses. Cela contraste avec l'approche de l'écologie comportementale, qui a donné naissance à une littérature de plus en plus abondante sur l'adaptation comportementale humaine. Dans cette revue, j'illustre de nombreux aspects de l'histoire de la vie humaine à partir de données provenant d'un village d'Afrique de l'Ouest, recueillies entre 1950 et 1975. Cet ensemble de données constitue l'un des registres les plus complets de la démographie d'une population humaine naturelle, aux ressources alimentaires limitées. Ces données ont été collectées en temps réel dans le cadre d'un projet de recherche médicale à long terme, mené dans un village gambien où la population dépendait de l'agriculture et souffrait d'une mortalité élevée. J'ai choisi cette population pour illustrer une vision écologique évolutionniste de l'histoire de la vie humaine, non pas parce qu'elle prétend être ancestrale, mais simplement parce qu'il s'agit d'une population traditionnelle bien documentée, aussi susceptible d'être caractéristique de l'humanité que n'importe quelle autre.
Compromis d’histoire de vie
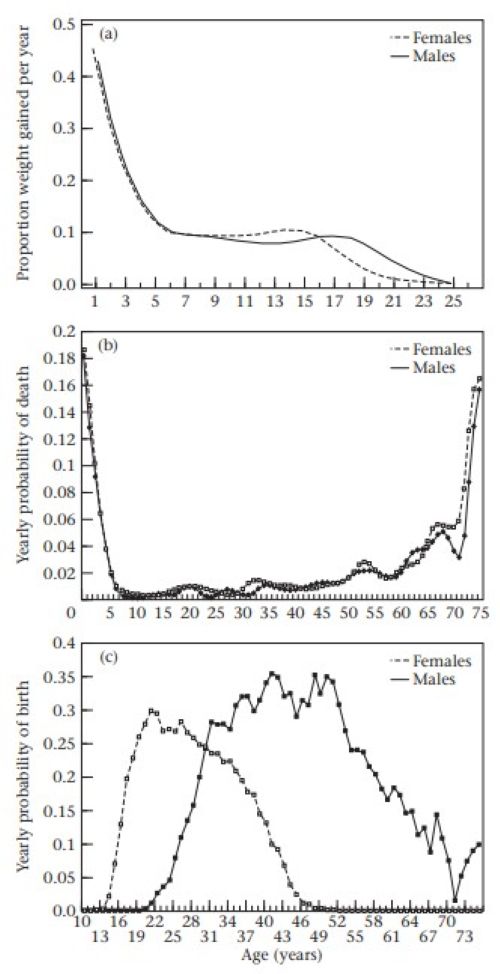
La figure 1 illustre les caractéristiques essentielles du cycle biologique humain : croissance, mortalité et fertilité chez des Gambiens ruraux entre 1950 et 1975.
(a) Gain de poids annuel moyen en proportion du poids total, entre la naissance et 25 ans, pour les hommes et les femmes.
(b) Risque de mortalité annuel pour les hommes et les femmes tout au long de la vie.
(c) Fécondité par âge (nombre de naissances vivantes par an) pour les hommes et les femmes tout au long de la vie (moyennes mobiles sur 3 ans).
Une hypothèse fondamentale de la théorie du cycle biologique est l’existence de compromis entre l’énergie dépensée pour la croissance et les facteurs influençant la mortalité, d’une part, et la reproduction, d’autre part[10],[11],[12]. Les coûts de la reproduction détournent l’énergie de la réparation et de l’entretien de l’organisme[13] et réduisent l’investissement dans la compétence immunologique. Des preuves de compromis entre l’effort reproductif et la durée de vie ont été recueillies grâce à des manipulations expérimentales chez de nombreuses espèces animales[14]. Un problème évident lié à l’obtention de telles preuves chez l’homme est que les chercheurs ont dû s’appuyer sur des corrélations phénotypiques ou des « expériences naturelles ». Les corrélations phénotypiques sont problématiques, car l'hétérogénéité d'une population peut masquer les véritables relations entre les variables du cycle biologique.
Par exemple, si les femmes se reproduisent au maximum de leurs capacités, les femmes en meilleure santé ont des familles plus nombreuses et potentiellement une plus grande longévité. En revanche, si cette reproduction supplémentaire réduit l'espérance de vie, aucune corrélation entre la taille de la famille et la longévité n'apparaît.
Malgré ces problèmes, des coûts immédiats, tant en termes de risque de mortalité maternelle que de stress nutritionnel après la reproduction, ont été démontrés dans les populations mal nourries[15]. Par exemple une étude a constaté que les femmes norvégiennes ayant moins de quatre enfants vivent plus longtemps que celles ayant plus de quatre enfants[16]. Une autre a montré qu'une procréation plus rapide chez les femmes de l'aristocratie anglaise était associée à une espérance de vie plus courte[17]. Une autre a montré qu'une reproduction tardive était associée à une meilleure longévité[18]. Il y a donc de nombreuses preuves d'un compromis génétique entre reproduction et longévité, cependant ces effets sont plutôt subtils et il n'est pas certain qu'ils agissent à des niveaux biologiquement significatifs.
En ce qui concerne l'évaluation du rôle de la fonction immunitaire dans le cycle biologique, la plupart des recherches pertinentes sur l'homme ont suivi une direction différente de celles menées sur d'autres animaux. Cela est dû à la disponibilité de données documentant les variations de la nutrition fœtale, fournissant des expériences naturelles qui semblent montrer que la gestation est une période critique pour la programmation de la fonction immunitaire plus tard dans la vie.
Une étude a montré qu'une mauvaise nutrition in utero, causée soit par de graves pénuries alimentaires pour les mères, soit par des complications médicales pendant la grossesse, est associée à des bébés maigres et à un risque considérablement accru de maladies dégénératives à l'âge adulte (comme le diabète et les crises cardiaques)[19]. Une étude a utilisé la saisonnalité pour montrer que les mères au troisième trimestre de grossesse pendant la saison peu nutritive, en Gambie, avaient des enfants plus susceptibles de mourir de maladies infectieuses à l'âge adulte[20]. Il reste à établir si le détournement d'énergie du système immunitaire à l'âge adulte influence les coûts de la reproduction.
Quels que soient les mécanismes précis des coûts de la reproduction, les femmes du monde entier en sont certainement bien conscientes. Hill en 1997 a relaté comment les Gambiennes décrivent les coûts de la reproduction tels qu'elles les perçoivent, à travers le concept mandingue de faaso : l'idée fondamentale est que les femmes possèdent une certaine capacité à la naissance ou au mariage, qui diminue progressivement au fil des événements reproductifs, et qu'une fois épuisée, elle ne peut être totalement reconstituée, même si une partie peut être reconstituée par le repos et une bonne alimentation… Une femme avec peu de faaso est « vieille ». Très peu d'hommes souhaitent des femmes ayant de faibles réserves de faaso. Il peut donc être prudent pour une femme, si son mariage est instable ou si son mari est malade, de conserver un peu de faaso en prévision d'un nouveau mariage imminent.
Sociobiologie pure. C'est l'utilisation optimale des facteurs tout au long de la vie qui constitue le défi de l'histoire de vie humaine.
Des compromis interviennent à chaque étape : entre croissance et reproduction, entre un enfant et un autre, entre enfants et petits-enfants. La théorie de l'histoire de vie est le seul paradigme théorique capable de prédire comment la sélection naturelle devrait résoudre ces compromis.
Le parcours de vie
Naissance et petite enfance
L'accouchement est difficile et douloureux chez l'humain, car le bébé est gros et notre posture verticale nécessite un bassin rigide. Chez la majorité des mammifères, la croissance cérébrale ralentit à la naissance, mais chez l'humain, le crâne n'est pas complètement durci à la naissance, afin de faciliter l'accouchement. Le cerveau se développe rapidement après la naissance pendant environ un an, atteignant la taille adulte en sept ans environ[21].
L'accouchement reste risqué, tant pour la mère que pour l'enfant. La mortalité maternelle varie énormément à travers le monde, atteignant un pic dans les hôpitaux où l'hygiène est déficiente, où l'accouchement peut être plus dangereux qu'à domicile. Dans les pays dépourvus de médecine moderne efficace, le risque de décès lors d'un accouchement hors hôpital est inférieur à 1 % par naissance, mais suit une courbe en J : il diminue après la première naissance, puis augmente à nouveau chez les femmes à parité élevée. Le taux global est peut-être inférieur à ce que notre peur de l'accouchement pourrait laisser penser, mais, la mortalité adulte étant faible chez l'homme, un risque de mortalité maternelle inférieur à 1 % par naissance se traduit généralement par entre 0,25 et 0,33 décès de femmes en âge de procréer dans les populations à fécondité naturelle comme l’Afrique[22]. La mortalité féminine est souvent supérieure à celle des hommes entre 20 et 30 ans, pour cette raison. La mise à disposition d'antibiotiques et de structures permettant une césarienne sécurisée réduit considérablement ce risque[23], et lorsque ces dispositifs sont disponibles, la mortalité maternelle est négligeable. Les risques pour le bébé sont bien plus élevés. Sauf avec les meilleurs soins médicaux, le risque de mortalité périnatale descend rarement en dessous de 1 %. Une partie de cette mortalité peut être due à des anomalies génétiques, qui ne sont fatales qu'après la naissance. L'exemple gambien de la figure 1 illustre l'extrême opposé : 7 % des nourrissons nés vivants décèdent dans le mois qui suit. Le poids corporel est un corrélat fort et constant des chances de survie d'un nourrisson au-delà de sa première année. Le sevrage est une période dangereuse, car les nourrissons sont exposés à des aliments moins nutritifs et à une eau insalubre. Plus le sevrage est précoce, plus la mortalité infantile est élevée. La raison la plus fréquente du sevrage est une grossesse ultérieure ; et les preuves que les naissances rapprochées augmentent considérablement la mortalité infantile sont accablantes[24],[25],[26] .Les frères et sœurs aînés se disputent alors la nourriture[27]
Les bébés humains sont également exposés au risque d'infanticide, ou de négligence entraînant une réduction de leur survie. Le scénario classique, décrit pour de nombreux mammifères, d'infanticide commis par les nouveaux mâles, afin de réorienter les efforts parentaux de la mère vers la future progéniture des nouveaux mâles, constitue un risque sérieux pour les enfants de moins de deux ans dans notre société[28]. Voland en 1988 a trouvé des preuves statistiques provenant de l'Allemagne du XVIIIe siècle indiquant que les femmes veuves, jeunes, pouvaient réduire les chances de survie de leurs jeunes enfants afin d'améliorer leurs chances de remariage. Un phénomène similaire se produit également chez nous[29]. L'abandon de la mère par son mari est fréquemment associé à l'infanticide, souvent à la naissance (par exemple, chez les Aché)[30]. Les préférences sexuelles peuvent également être à l'origine d'importants biais sexuels dans la mortalité infantile et juvénile. Ces préjugés peuvent être défavorables aux filles (comme c'est souvent le cas dans les sociétés patrilinéaires, notamment en Asie) ou aux garçons (notamment dans les sociétés matrilinéaires, comme en Afrique australe[31]). Les enfants du sexe le moins favorisé qui ont des frères et sœurs aînés du même sexe sont particulièrement à risque[32]. La majorité de ces tendances, qui indiquent l'influence des parents sur la mortalité infantile, ont été identifiées par une analyse statistique minutieuse des données démographiques plutôt que par des entretiens avec les parents. Cependant, lorsque les conditions économiques et le niveau de soutien familial et paternel sont très faibles, les mères admettent parfois leur réticence à se soumettre à l'infanticide par négligence, comme constaté dans les bidonvilles brésiliens[33].
Le bébé ne peut faire face aux aléas de la petite enfance, si ce n'est grandir le plus vite possible (Fig. 1a). À mesure qu'ils grandissent, les enfants nécessitent moins d'efforts parentaux et, s'échappant dans l'oasis de l'enfance, ils semblent généralement moins exposés au risque de mortalité (Fig. 1b).
Enfance et puberté
L'enfance n'est pas propre à l'homme, un phénomène similaire étant observé chez d'autres singes[34], mais elle est particulièrement prolongée chez l'homme. On considère généralement que l'enfance est liée au développement intellectuel et social plutôt qu'au développement physique, bien que certains s'opposent à cette vision. L'absence de preuves tangibles que ce que nous apprenons pendant l'enfance contribue à notre condition physique est flagrante. Le rattrapage de croissance, observé chez les enfants se remettant d'une maladie ou d'une période de malnutrition, illustre qu'une croissance bien plus rapide que celle observée est physiologiquement possible. Cependant, l'idée d'un enfant de 5 ans, adulte et pleinement développé, est effrayante, et il est peu probable qu'il réussisse particulièrement bien dans la compétition pour les ressources ou les partenaires avec des jeunes de 20 ans. Il est préférable qu'une période d'apprentissage, ou simplement d'attente, soit préservée grâce à des besoins nutritionnels moindres, une menace moindre pour les adultes et une faible mortalité. Dans le même ordre d'idées, Haig a décrit l'enfance comme une stratégie visant à obtenir un plus grand investissement parental[35].
À la puberté, les enfants connaissent une poussée de croissance rapide jusqu'à atteindre ou presque leur taille maximale. Cette poussée de croissance à l'adolescence est une spécificité humaine.
Les poussées de croissance sont plus prononcées dans les populations bien nourries que dans les populations mal nourries, comme dans le cas gambien. Dans toutes les populations étudiées, les hommes atteignent une taille adulte moyenne supérieure à celle des femmes, bien que la variation interpopulationnelle du dimorphisme sexuel soit probablement liée à l'investissement parental biaisé en fonction du sexe pendant l'enfance[36].
Les poussées de croissance féminines surviennent à un âge plus précoce que celles des hommes et ont tendance à survenir avant que l'enfant ne devienne fertile, peut-être en raison des graves conséquences d'une grossesse pendant le sous-développement des filles. En particulier, la croissance de la largeur des hanches est associée à l'apparition de la fertilité[37]. Une maturation précoce peut également être dangereuse pour les garçons : les garçons Ache sont initiés dès qu'ils commencent à faire des avances sérieuses aux filles, ce qui en fait des cibles potentielles dans des bagarres avec des hommes adultes[38].
Le début de la puberté, qui coïncide avec le ralentissement de la croissance chez les filles, semble présenter une grande plasticité en fonction des circonstances environnementales. Il n'existe ni taille ni âge fixes à la ménarche, mais une norme réactionnelle, selon laquelle les filles malnutries atteindront leurs ménarches plus tard, mais à une taille plus petite[39]. Une tendance séculaire à Une puberté plus précoce a été observée partout dans les populations bien nourries.
Accouplement et mariage
Dans les sociétés humaines, les schémas d'accouplement sont fréquemment légalisés par le mariage, et les schémas matrimoniaux sont très diversifiés. La majorité des cultures présentent une résidence matrimoniale patrilocale. Les données génétiques montrent que la variabilité spatiale des chromosomes Y semble moindre que celle de l'ADNmt ou des chromosomes X, confirmant que nous sommes essentiellement patrilocaux, c'est-à-dire que les femmes sont plus susceptibles de migrer[40]. Nos plus proches parents primates, les chimpanzés (Pan troglodytes) et les bonobos (Pan paniscus), présentent une dispersion des femelles, ce qui a été utilisé pour affirmer que la patrilocalité est ancestrale chez l'homme ; cependant, compte tenu de 5 millions d'années d'évolution et de la diversité des schémas matrimoniaux au sein de notre espèce, nous ne devrions pas considérer cela comme une certitude. Les chasseurs-cueilleurs présentent une lignée diversifiée, voire inexistante ; et l’origine des richesses défendables et héréditaires change tout.
Cependant, une minorité significative de groupes humains présente une descendance matrilinéaire, où les richesses (comme la terre) sont transmises par la lignée féminine, et les mères vivent avec leurs filles, qui héritent de leurs champs ; la résidence conjugale est matrilocale. Les hommes possèdent peu de biens personnels, mais sont censés les transmettre aux enfants de leurs sœurs, et non aux leurs[41]. Les mariages sont généralement instables ; l’investissement d’un homme dans les enfants de sa femme étant faible, l’importance accordée à la fidélité conjugale est faible.
Avec l’émergence de richesses précieuses et héréditaires, comme le bétail, qui doit être protégé contre le vol par les hommes, la patrilinéarité est devenue majoritaire[42],[43],[44]. Ici, la richesse se transmet de père en fils, la résidence conjugale est patrilocale et l'accent est mis sur la fidélité conjugale. Les épouses, souvent achetées avec une dot, deviennent des biens qui ne sont pas libres de partir, car elles perdraient richesse et enfants.
On ne sait pas clairement dans quelle mesure les schémas matrilinéaires reflètent les schémas d'accouplement. Il est difficile d'obtenir des données directes sur l'incertitude de paternité chez l'homme pour des raisons éthiques, mais plusieurs estimations se situent autour de 10 %. Une estimation de l'incertitude de paternité a utilisé la théorie de la sélection de parentèle de Hamilton pour montrer que les parents matrilinéaires investissent davantage dans leurs enfants que les parents patrilinéaires en Europe occidentale et aux États-Unis, dans des proportions compatibles avec une incertitude de paternité comprise entre 9 et 15 %[45],[46]. Les attributs physiques considérés comme attrayants chez les partenaires ont été largement étudiés ailleurs, et je ne les développerai pas ici, mais deux influences claires et constantes sont l'âge et la richesse. Les différences de fécondité entre les sexes tout au long de la vie montrent clairement que les femmes sont les plus recherchées comme partenaires de mariage lorsqu'elles sont jeunes, mais que les hommes peuvent atteindre une fécondité élevée même à un âge avancé (Fig. 1c). La mesure dans laquelle cela est vrai dépend encore une fois du contexte socio-économique dans lequel les individus vivent. Le mariage polygyne et la propriété patrilinéaire des ressources signifient que les hommes âgés peuvent épouser des jeunes femmes s'ils possèdent les ressources nécessaires pour subvenir à leurs besoins et à ceux de leurs enfants. Il peut être avantageux pour les jeunes femmes d'épouser des hommes polygames plus âgés s'ils sont plus aisés. Une étude a utilisé des données démographiques historiques pour montrer que les épouses mormones polygames de l'Utah au XVIIIe siècle avaient plus de petits-enfants que les femmes mariées monogames[47]. Lorsque les ressources héritées contribuent fortement au succès reproductif futur, le mariage peut être retardé et la compétition entre frères et sœurs peut réduire directement le succès reproductif de ceux qui héritent moins[48],[49].
Le rythme de la procréation
Le calendrier de la reproduction chez les femmes présente des caractéristiques remarquablement différentes de celui des grands singes. Premièrement, les intervalles entre les naissances de 2,5 à 3,5 ans semblent plutôt courts compte tenu de notre taille (par exemple, les gibbons hylobates ont des intervalles entre les naissances de 3 ans, les chimpanzés de 4 à 5 ans, et les orangs-outans de près de 8 ans). Deuxièmement, la reproduction s'arrête vers la moitié de l'âge adulte avec la ménopause, une sénescence programmée des organes reproducteurs peut-être de plus de 20 ans avant la sénescence du reste du corps. La capacité biologique des femmes à se reproduire après chaque naissance est dictée par plusieurs facteurs énergétiques qui influencent l'ovulation, notamment les habitudes d'allaitement et la charge de travail[50]. La grande majorité des naissances sont des naissances simples, bien que les taux de gémellité augmentent avec l'âge et persistent à des niveaux suffisamment élevés pour que la gémellité ait pu être sélectionnée dans certaines circonstances, malgré une mortalité gémellaire élevée. Une étude a calculé que, dans l'Allemagne du XVIIIe siècle, la gémellité n'entraînait une augmentation de la fitness à long terme (nombre de petits-enfants) que si les deux jumeaux étaient des filles[51]. Cependant, les mères de jumeaux sont souvent particulièrement fertiles et la gémellité pourrait être maintenue comme un effet secondaire pléiotrope de ce trait bénéfique[52].
L'intervalle des naissances tend à augmenter avec l'âge, même avant la ménopause. Cela peut être lié au vieillissement ou à une réaction évolutive liée à la compétition entre frères et sœurs. Certaines sociétés ont démontré qu'un ralentissement de la reproduction est associé à l'obtention de la taille de famille souhaitée. Par exemple, l'absence de fils dans une société patrilinéaire est associée à une reproduction continue[53],[54]. Le rythme relativement rapide des grossesses au cours de la première moitié de l'âge adulte conduit à la naissance d'enfants à différents stades de dépendance, nécessitant des soins simultanés. La possibilité de fonder une telle famille a été attribuée à la capacité des mères à s'assurer l'aide d'autres membres de la famille pour les soins et l'alimentation des enfants. L'apport des mâles, absent chez les autres primates, pourrait permettre des taux de natalité élevés chez les femelles humaines[55], mais les femelles ménopausées pourraient également être à l'origine de cette contribution énergétique supplémentaire à la famille[56]. Il n'y a pas de consensus, mais il semble peu probable que l'éducation d'une famille humaine ait jamais été l'affaire des seules mères. La garde des enfants était peut-être une responsabilité pour tous, sauf pour les plus jeunes, qu'il s'agisse d'un aîné, d'un parent ou d'un grand-parent. Avec la dispersion de la famille élargie à l'ère moderne, la parentalité devient une période mouvementée de la vie, rythmée de part et d'autre par de longues périodes d'activités éducatives et récréatives de plus en plus élaborées, ou par des moments de va-et-vient.
L'évolution de la ménopause
La ménopause a peut-être évolué précisément pour que les femmes âgées puissent aider leurs enfants à se reproduire plutôt que de continuer à le faire elles-mêmes, une théorie connue sous le nom d'hypothèse des grands-mères[57],[58]. Si les femmes se sont dispersées au moment du mariage, il pourrait être nécessaire d'avancer que les grands-mères aidaient leurs belles-filles plutôt que leurs filles. Le lien génétique entre l'enfant d'une belle-fille et l'enfant d'une fille diffère selon le degré d'incertitude de la paternité, et l'idée d'une belle-mère bienveillante semble un peu moins vraie, par exemple, le décès prématuré d'une belle-mère augmente l'espérance de vie des femmes japonaises[59]. Ainsi, la question de l'évolution du rythme des grossesses et celle de la ménopause sont liées, ainsi qu'à la question de la structure de notre famille ancestrale.
Les preuves que les femmes peuvent augmenter le nombre de leurs petits-enfants suffisamment pour compenser la perte de leurs propres possibilités de procréation sont mitigées. Des modèles simples ne parviennent pas à montrer que le renoncement à la reproduction au profit du fait d’être grand-mère comme évolutivement stable[60],[61].
Cependant, ces modèles reposent sur l'hypothèse que les risques liés à la reproduction et les avantages d’être grand-mère doivent l'emporter sur les avantages d'une reproduction continue au rythme d'une jeune femme, ceci ne tient pas compte d’un large éventail de coûts et d'avantages qui évoluent avec l'âge. Dans un modèle de cycle biologique féminin incluent la sénescence biologique généralisée, les critères favorisant la ménopause apparaissent beaucoup moins stricts[62]. Le risque de mortalité maternelle est un paramètre clé dans tous ces modèles, car c'est lui qui priverait les enfants existants de soins maternels si la fertilité se poursuivait jusqu'à un âge avancé. Le risque de mortalité maternelle a généralement été considéré comme constant, mais en réalité, il augmente de façon exponentielle avec l'âge, tout comme la mortalité néonatale[63].
Les données des maternités des États-Unis et du Royaume-Uni dans les années 1930 montrent que les causes de mortalité maternelle qui augmentent avec l'âge n'étaient pas liées à une infection, mais à une hémorragie et à un placenta mal positionné, ce qui pourrait suggérer une diminution de la force et du tonus musculaires plutôt qu'une diminution de l'immunocompétence. Chez les femmes âgées, l'utérus est incapable de se contracter pour prévenir les pertes sanguines aussi efficacement que chez les femmes plus jeunes, un problème désormais anticipé grâce aux injections d'ocytocine synthétique juste après l'accouchement. Il est possible que la combinaison de nos gros bébés et du rythme de reproduction rapide que notre système social autorise présente des risques très élevés pour la poursuite de la reproduction jusqu'à un âge avancé. Si ces risques venaient à s'accélérer encore avec l'âge, ce qui n'est pas improbable compte tenu de la nature multifactorielle du vieillissement. Survivre à un accouchement naturel à un âge avancé pourrait devenir hautement improbable et l'arrêt de la reproduction à 50 ans ne nécessiterait pratiquement aucune autre explication.
Notre période de reproduction n'est pas particulièrement courte, car elle correspond à un modèle plus général chez les primates[64], donc une longue durée de vie postménopausique est le seul caractère dérivé qui nécessite une explication[65]. Si cela est vrai, les bénéfices de la grand-mère en termes de fitness ne semblent pas si importants, bien que cet argument n'explique pas pourquoi les hommes (sans ménopause) ont généralement une longévité similaire à celle des femmes. Les enfants Hadza (cueilleurs) qui ont un parent ménopausé dans leur groupe familial sont mieux nourris que ceux qui n'en ont pas. La contribution de la grand-mère semble particulièrement importante lorsque la mère a un autre enfant. Les tubercules, difficiles à extraire et à transformer pour les enfants, semblent bien plus importants dans l'alimentation que la viande. Des auteurs ont avancé que l'aide grand-mère pourrait être responsable du sevrage précoce chez l'homme[66].
Outre le risque de mortalité maternelle évoqué précédemment, plusieurs raisons, générales et spécifiquement humaines, justifient que les familles ne soient pas trop nombreuses. Dans toutes les sociétés, seule une faible proportion des naissances concerne des femmes de plus de 40 ans, même si la ménopause est plus proche de 50 ans[67].
La ménopause serait beaucoup plus avantageuse si les autres coûts liés à l’existence d’autres enfants étaient inclus. La concurrence entre frères et sœurs pour l'investissement parental est l'un de ces facteurs. Bien qu'il ne soit pas certain que les effets de la compétition entre frères et sœurs sur la mortalité soient suffisants pour freiner la reproduction, elle pourrait néanmoins avoir quelques effets sur le succès reproductif futur[68],[69]. J'ai modélisé un calendrier reproductif optimal lorsque les frères et sœurs se disputent la richesse parentale, cette richesse influence leur succès reproductif futur. Les modèles montrent qu'il est très rarement optimal pour les femmes d'avoir une fertilité maximale ; ainsi, dans un large éventail de cas réalistes, le coût de la ménopause est pratiquement nul. La compétition pour la nourriture, le statut, les territoires, les opportunités de reproduction, le soutien familial ou le faaso pourrait suivre une dynamique similaire, ce qui entraînerait des rendements décroissants pour les familles nombreuses. Ainsi, seuls de faibles coûts biologiques liés au maintien de la fertilité féminine au-delà de 50 ans pourraient favoriser la ménopause.
Dans de telles circonstances, les bienfaits de la sollicitude parentale et grand-parentale n’ont pas à être si importants pour prolonger la vie. Et la liste des bienfaits possibles est longue. Une mère vivante semble être associée à un âge de première reproduction avancé d'environ un an chez ses filles, et les bébés ont plus de chances de survivre si leur grand-mère maternelle est vivante. Cette analyse préliminaire de la survie des premiers-nés suggère que les bienfaits d'une grand-mère maternelle vivante apparaissent après les 18 premiers mois, ce qui est cohérente avec l'hypothèse selon laquelle ces grands-mères contribuent également à l'alimentation de leurs petits-enfants après le sevrage.
Les femmes mariées ont tendance à vivre avec des parents patrilinéaires dans ce cas, mais généralement pas très loin de leur domicile maternel. Les bénéfices à long terme d'une reproduction précoce chez les filles pourraient être encore plus importants, bénéficiant à tous leurs futurs enfants et petits-enfants, ce qui rend les bénéfices des soins aux filles existantes potentiellement supérieurs aux bénéfices d’avoir des bébés plus tard dans la vie.
La transition démographique
Le changement le plus profond de l'histoire de l'humanité s'est produit récemment, et il est culturel. Il y a environ deux siècles, pendant une période d'expansion démographique très rapide, la fécondité en Europe et en Amérique du Nord a commencé à baisser, tout comme la mortalité. Ce phénomène s'est produit beaucoup plus tard en Asie et en Amérique du Sud, et a récemment commencé en Afrique[70]. La progression de ce phénomène démographique et l’épidémiologie conséquente ne sont pas les mêmes dans chaque région, mais il existe suffisamment de similitudes à travers le monde pour décrire ce phénomène et ses conséquences. Ceci intéresse particulièrement les anthropologues évolutionnistes, car une réduction de la taille des familles en période d'abondance des ressources est paradoxale sans le cadre d’une vision adaptative du comportement et du cycle biologique humains.
Le temps écoulé est insuffisant pour nous être adapté génétiquement à certains aspects de notre cycle biologique moderne. Notre faible taux de reproduction, tout en nous soulageant de certains problèmes médicaux anciens, nous en présente de nouveaux, dont possiblement les cancers de l'appareil reproducteur. Or, les décisions en matière de reproduction sont malléables dans les sociétés traditionnelles et modernes, et il est donc intéressant d'examiner comment l’évolution psychologique a pu produire une tendance aussi constante vers les familles de petite taille.
Les écologistes évolutionnistes connaissent bien le fait que la fertilité pourrait être réduite par un compromis quantité/qualité[71]. Les populations homogènes étudiées classiquement présentent fréquemment une corrélation positive entre richesse et succès reproductif, comme le prédit la théorie de l'écologie évolutionniste[72],[73]. Alors que les grandes populations hétérogènes étudiées par les démographes présentent généralement une corrélation négative entre richesse et succès reproductif. J'ai exploré ce paradoxe en utilisant un modèle pour prédire à la fois la taille optimale de la famille et le montant optimal de richesse à allouer à chaque enfant à la fin de la vie reproductive des parents, lorsque la richesse héritée des parents était considérée comme un déterminant important du succès reproductif futur[74]. Les deux variables sont liées, car la richesse consacrée à l'éducation des enfants n'est pas disponible pour leur héritage. Le fait important que la richesse elle-même puisse générer davantage de richesse a été intégré au modèle. L'optimalité a été définie comme la maximisation du nombre de petits-enfants. Trois variables fréquemment associées aux transitions démographiques ont été modifiées dans le modèle : le risque de mortalité extrinsèque, le risque de sécheresse ou de pénurie de ressources et le coût de l’éducation des enfants. Le modèle a montré que le risque de mortalité, bien qu’ayant une forte influence sur le nombre de naissances, n’avait pratiquement aucun effet sur la taille optimale de la famille ni sur le montant du patrimoine alloué à chaque enfant. L’atténuation du risque de sécheresse prédisait une augmentation de la taille de la famille et réduisait les risques sur l’héritage, appauvrissant les famille de taille moyenne. L'augmentation du coût de l'éducation des enfants était la seule variable produisant des résultats présentant les caractéristiques d'une transition démographique : la taille optimale des familles diminuait, et la richesse optimale allouée à chaque enfant augmentait, enrichissant les familles de taille moyenne.
Ces résultats concordent avec les données sur les réponses démographiques immédiates des personnes confrontées aux changements environnementaux. Des soins médicaux gratuits sont disponibles dans le village gambien de Keneba depuis 1975, ce qui a considérablement réduit la mortalité, mais rien n'indique le début d'une transition démographique. Ceci est à comparer avec des exemples de transition démographique quasi instantanée lorsque les variations de la fécondité changent vite lors de modifications des coûts et avantages économiques liés à l'éducation des enfants[75],[76].
Le modèle permet de comprendre pourquoi une corrélation négative entre richesse et succès reproductif est fréquemment observée. Au sein de chaque population modèle, la richesse était positivement associée au succès reproductif, mais paradoxalement, les populations modèles dont les familles étaient les plus pauvres étaient également celles dont la taille était la plus importante. Si des populations hétérogènes représentent des sous-populations, chacune subissant (ou croyant subir) des coûts et des avantages différents liés à l'investissement dans les enfants, alors un découplage ou une relation négative entre richesse et taille de la famille pourrait être observé. Une étude dans une société homogène post-transition de femmes britanniques diplômées des universités anglaises dans les années 1930 et 1940 a montré que la richesse était positivement corrélée au succès reproductif[77]. Le niveau de diplôme obtenu par ces femmes était également corrélé positivement à leur fécondité future. Ce dernier résultat est intrigant, car le niveau d'éducation est généralement associé à une faible fécondité ; mais, là encore, il pourrait s'agir d'une confusion entre les réponses interpopulationnelles et individuelles. Kaplan a développé en 1996 le concept de « capital incorporé », un terme utilisé pour englober tous les bénéfices que les parents peuvent investir dans leurs enfants et qui pourraient se traduire par des effets à long terme. Ces bénéfices peuvent concerner la réussite scolaire, la capacité de gain, la santé ou la fécondité. Si notre psychologie évoluée peut favoriser l'amélioration de la qualité de vie de notre progéniture, il n'est pas certain que cela se traduise par une réussite reproductive dans le monde moderne. Dans un échantillon contemporain d'hommes du Nouveau-Mexique, ceux nés dans des familles plus petites avaient de meilleurs résultats scolaires et une meilleure capacité de gain, mais cela ne se traduisait pas par une meilleure fitness[78]. Cependant, ces données couvraient une période d’expansion économique et démographique : ce capital incorporé supplémentaire aurait pu générer de meilleurs rendements en termes de fitness dans une population stable, où la concurrence est plus intense.
La majorité des écologistes évolutionnistes ne sont pas en mesure d'étudier le processus historique de changement comportemental des populations, contrairement à ce que font souvent les écologistes évolutionnistes humains. Cela nécessite d'étudier les mécanismes associés à l'adaptation et leur évolution temporelle. À cet égard, des études en psychologie, en économie et dans d'autres sciences sociales, qui dépassent le cadre de cette revue, recoupent des études sur la fertilité humaine. Cependant, le début de la transition démographique ne peut être systématiquement rattaché à une variable biologique, écologique ou économique particulière. Il se peut que la notion d'investissement parental élevé dans chaque enfant soit considérée comme un « même » (au sens de Dawkins) qui se répand dans le monde entier, et qui est particulièrement efficace dans certaines circonstances. Une analyse minutieuse de la baisse de la fertilité en France, pays qui a connu la première et la plus connue des transitions démographiques, montre comment les familles plus petites, vraisemblablement associées à un investissement parental plus élevé dans chaque enfant, se sont propagées au fil du temps à partir de centres de sophistication[79]. Dans les régions reculées, la baisse de la fertilité est survenue plus tard, mais une fois amorcée, elle s'est poursuivie plus rapidement. Aucun corrélat socio-économique simple ne prédit sa propagation, si ce n'est peut-être l'éloignement des épicentres.
La compétition doit être essentielle. Il est certainement intéressant psychologiquement, d'investir dans nos enfants afin qu'ils puissent rivaliser avec leurs pairs, sans doute pour de bonnes raisons évolutionniste. Les enfants des villages finissent par rivaliser avec ceux d'autres villages, d'autres villes, voire d'autres pays, à mesure que les populations deviennent plus mobiles ; ils se disputent les résultats aux examens, les emplois et les partenaires. Lorsqu'une partie de la population investit massivement dans sa progéniture, d'autres suivront. Je viens de mettre à niveau mon ordinateur pour que mon enfant de 4 ans puisse maintenir des compétences informatiques comparables à celles des autres enfants de sa crèche, et je peux témoigner que ce processus commence tôt. Lorsque les coûts sont envisagés en termes de compétition entre enfants plutôt que de simples coûts de subsistance, il est possible d'imaginer que nous ne sommes pas aussi éloignés des contraintes de ressources sur la reproduction que nos modes de vie le laissent entendre. Lorsque la richesse est clairement abondante, le temps peut être la ressource que les parents ne peuvent pas allouer à leurs enfants en quantités optimales.
Conclusion
J'ai tenté d'illustrer comment les approches issues de l'écologie évolutionniste contribuent à la compréhension de nos propres vies. Bien qu'il soit tentant de relier tous les étranges attributs du cycle biologique humain à un seul effet (comme notre cerveau puissant, notre faible mortalité adulte, ou encore un aspect de notre système social ou de notre alimentation), l'un des enseignements des études sur le cycle biologique est que différents traits sont soumis à des pressions de sélection différentes. C'est la combinaison des pressions de sélection influençant la mortalité et la fertilité qui favorise une stratégie de cycle biologique particulière, et les modèles révèlent que de faibles variations de ces pressions de sélection peuvent avoir des effets importants sur le cycle biologique optimal. Même chez des espèces étroitement apparentées, la différence des compromis semble s'avérer importante. D'autres études révéleront les variations interculturelles du cycle biologique humain et les compromis observés. Avec le temps, nous pourrons voir si des tendances cohérentes se dégagent.
Les écologistes évolutionnistes se sont intéressés tardivement à l'étude de la fertilité, de la mortalité et de la croissance humaines. Médecins, anthropologues sociaux, psychologues, démographes et économistes ont été les premiers à intervenir et n'ont pas toujours accueilli favorablement les nouveaux venus. Cependant, ils n'ont pas résolu, ni même identifié, tous les problèmes intéressants posés par l'évolution de l'histoire de la vie humaine. Le paradigme de l'écologie évolutionniste apporte une nouvelle perspective, de nouveaux outils et de nouveaux chercheurs pour cette tâche inachevée et colossale.
Traduction par Luc Perino de l'article :
Mace R. Evolutionary ecology of human life history. Anim Behav. 2000 Jan;59(1):1-10. DOI: 10.1006/anbe.1999.1287
Références
[1] Pagel & Harvey, 1989
[2] Howell, 1979
[3] Pennington, 1992
[4] Hill et Hurtado,1996
[5] Symons 1979
[6] Simoons, 1978
[7] Holden et Mace, 1997
[8] Strassman et Dunbar, 1999
[9] Boyd et Richerson, 1985
[10] Williams 1957
[11] Roff 1992
[12] Stearns, 1992
[13] Kirkwood et Rose, 1991
[14] Roff, 1992
[15] Tracer, 1991
[16] Lund, 1990
[17] Westendorp et Kirkwood, 1998
[18] Perls, 1997
[19] Barker, 1994
[20] Moore, 1997
[21] Bogin, 1999
[22] Mace et Sera, 1996
[23] Graham, 1991
[24] Hobcraft, 1983
[25] Alam, 1995
[26] Bohler et Bergstrom, 1995
[27] Le Grand et Philip,s 1996
[28] Daly et Wilson, 1985
[29] Daly et Wilson, 1996
[30] Hill et Hurtado, 1996
[31] Harpending et Pennington, 1991
[32] DasGupta, 1987
[33] Scheper-Hughes, 1992
[34] Perreira et Fairbanks, 1991
[35] Haig, 1999
[36] Holden & Mace, 1999
[37] Bogin, 1999
[38] Hill et Hurtado, 1996
[39] Stearns et Koella, 1986
[40] Seielstad, 1998
[41] Schneider et Gough, 1961
[42] Engels, 1884
[43] Aberle, 1961
[44] Mace et Holden, 1999
[45] Gaulin, 1997
[46] Euler et Weitzel, 1996
[47] Josephson, 1993
[48] Johansson, 1987
[49] Low, 1991
[50] Ellison, 1994
[51] Gabler et Voland, 1994
[52] Lummaa , 1998
[53] Nath et Land, 1994
[54] Mace et Sear, 1997
[55] Hill, 1993
[56] Hawkes, 1997
[57] Williams, 1957
[58] Hamilton, 1966
[59] Skinner, 1997
[60] Hill & Hurtado 1991, 1996
[61] Rogers, 1993
[62] Shanley,1999
[63] Loudon, 1992
[64] Charnov, 1993
[65] Hawkes, 1997
[66] Blurton-Jones, 1999
[67] Wood, 1994
[68] Hill et Hurtado, 1996)
[69] Rogers, 1993
[70] Cleland, 1995
[71] Lack, 1968
[72] Irons, 1979
[73] Borgerhoff-Mulder, 1987
[74] Mace, 1998
[75] Hoem, 1992
[76] Conrad, 1996
[77] Hubback, 1957
[78] Kaplan, 1995
[79] Bonneuil, 1997
Bibliographie complète
- Aberle, D. F. 1961. Matrilineal descent in cross-cultural perspective. In: Matrilineal Kinship (Ed. by D. M. Schneider & K. Gough), pp. 655–730. Berkeley: University of California Press.
- Alam, M. 1995. Birth-spacing and infant and early childhood mortality in a high fertility area of Bangladesh: age-dependent and interactive effects. Journal of Biosocial Science, 27, 393–404.
- Barber, N. 1995. The evolutionary psychology of physical attractiveness: sexual selection and human morphology. Ethology and Sociobiology, 16, 395–424.
- Barker, D. J. P. 1994. Mothers, Babies and Disease in Later Life. London: BMJ Publishing.
- Blurton-Jones, N., Hawkes, K. & O’Connell, J. F. 1999. Some current ideas about evolution of the human life history. In: Comparative Primate Socioecology (Ed. by P. C. Lee), pp. 140–166. Cambridge: Cambridge University Press.
- Bogin, B. 1999. Patterns of Human Growth. 2nd edn. Cambridge: Cambridge University Press.
- Bohler, E. & Bergstrom, S. 1995. Subsequent pregnancy effects morbidity of previous child. Journal of Biosocial Sciences, 27, 431–442.
- Bonneuil, N. 1997. Transformation of the French Demographic Landscape 1806–1906. Oxford: Clarendon Press.
- Borgerhoff-Mulder, M. 1987. On cultural and biological success: Kipsigis evidence. American Anthropologist, 89, 619–634.
- Boyd, R. & Richerson, P. J. 1985. Culture and the Evolutionary Process. Chicago: University of Chicago Press.
- Buss, D. M. 1989. Sex differences in human mate preferences: evolutionary hypotheses testing in 37 cultures. Behavioural and Brain Sciences, 12, 1–49.
- Cavalli-Sforza, L. L., Menozzi, P. & Piazzo, A. 1994. The History and Geography of Human Genes. Princeton, New Jersey: Princeton University Press.
- Charnov, E. L. 1993. Life History Invariants. Oxford: Oxford University Press. Cleland, J. 1995. Obstacles to fertility decline in developing countries. In: Human Reproductive Decisions (Ed. by R. Dunbar), pp. 207–229. London: MacMillan.
- Conrad, C., Lechner, M. & Werner, W. 1996. East German fertility after unification: crisis or adaptation. Population Development Review, 22, 331–358.
- Daly, M. & Wilson, M. I. 1985. Child abuse and other risks of not living with both parents. Ethology and Sociobiology, 6, 155–176.
- Daly, M. & Wilson, M. I. 1996. Violence against stepchildren. Current Directions in Psychological Science, 5, 77–81.
- DasGupta, M. 1987. Selective discrimination against female children in rural Punjab. Population Development Review, 13, 77–100.
- Dawkins, R. 1976. The Selfish Gene. Oxford: Oxford University Press.
- Ellison, P. T. 1994. Advances in human reproductive ecology. Annual Review of Anthropology, 23, 255–275.
- Engels, F. 1884. The Origin of the Family, Private Property and the State. London: Penguin.
- Euler, H. A. & Weitzel, B. 1996. Discriminative grandparental solicitude as reproductive strategy. Human Nature, 7, 39–59.
- Gabler, S. & Voland, E. 1994. The fitness of twinning. Human Biology, 66, 699–713.
- Gaulin, S. J. C., McBurney, D. H. & Brakeman-Wartell, S. L. 1997. Matrilateral biases in the investment of aunts and uncles: a consequence and measure of paternity uncertainty. Human Nature, 8, 139–151.
- Graham, W. 1991. Maternal mortality: levels, trends and data deficiencies. In: Disease and Mortality in Sub-Saharan Africa (Ed. by R. Feacham & D. Jamison), pp. 101–126. Oxford: Oxford University Press.
- Haig, D. 1999. Genetic conflicts of pregnancy and childhood. In: Evolution in Health and Disease (Ed. by S. C. Stearns), pp. 77–89. Oxford: Oxford University Press.
- Hamilton, W. D. 1964. The genetical evolution of social behaviour. Journal of Theoretical Biology, 7, 1–52.
- Hamilton, W. D. 1966. The moulding of senescence by natural selection. Journal of Theoretical Biology, 12, 12–45.
- Harpending, H. C. & Pennington, R. 1991. Age structure and sex-biased mortality among Herero Pastoralists. Human Biology, 63, 329–353.
- Hawkes, K., O’Connell, J. F. & Blurton-Jones, N. G. 1997. Hadza women’s time allocation, offspring provisioning, and the evolution of long post-menopausal lifespans. Current Anthropology, 38, 551–578.
- Hill, A. 1997. Truth lies in the eye of the beholder: the nature of evidence in demography and anthropology. In: Anthropological Demography (Ed. by D. Kertzer & T. Fricke), pp. 223–247. Chicago: University of Chicago Press.
- Hill, K. 1993. Life history theory and evolutionary anthropology. Evolutionary Anthropology, 2, 78–88. Hill, K. & Hurtado, A. M. 1991. The evolution of reproductive senescence and menopause in human females. Human Nature, 2, 315–350.
- Hill, K. & Hurtado, A. M. 1996. Ache Life History: the Ecology and Demography of a Foraging People. New York: Aldine de Gruyter.
- Hobcraft, J., McDonald, J. W. & Rutstein, S. 1983. Child-spacing effects in infant and early child mortality. Population Index, 49, 585–618.
- Hoem, J. 1992. Social policy and recent fertility change in Sweden. Population Development Review, 16, 735–748.
- Holden, C. & Mace, R. 1997. A phylogenetic analysis of the evolution of lactose digestion in adults. Human Biology, 69, 605–628.
- Holden, C. & Mace, R. 1999. Sexual dimorphism in stature and women’s work: a cross-cultural analysis. American Journal of Physical Anthropology, 110, 27–45.
- Howell, N. 1979. Demography of the Dobe area !Kung. New York: Academic Press.
- Hubback, J. 1957. Wives Who Went to College. London: Heinemann. Irons, W. 1979. Cultural and biological success. In: Evolutionary Biology and Human Social Behaviour: an Anthropological Perspective (Ed. by N. Chagnon & W. Irons), pp. 257–272. North Scituate: Duxbury.
- Johansson, S. 1987. Status anxiety and demographic contraction of privileged populations. Population Development Review, 13, 439– 470.
- Josephson, S. C. 1993. Status, reproductive success and marrying polygynously. Ethology and Sociobiology, 14, 391–396.
- Kaplan, H. 1996. A theory of fertility and parental investment in traditional and modern human societies. Yearbook of Physical Anthropology, 39, 91–135.
- Kaplan, H., Lancaster, J., Bock, J. A. & Johnson, S. E. 1995. Does observed fertility maximise fitness among New Mexican men? A test of an optimality model and a new theory of parental investment in the embodied capital of offspring. Human Nature, 6, 325–360.
- Kirkwood, T. B. L. & Rose, M. R. 1991. Evolution of senescence: late survival sacrificed for reproduction. Philosophical Transactions of the Royal Society of London, Series B, 332, 15–24.
- Lack, D. 1968. Ecological Adaptations for Breeding in Birds. London: Methuen.
- LeGrand, T. & Phillips, J. F. 1996. The effect of fertility reductions on infant and child mortality: evidence from Matlab in rural Bangladesh. Population Studies, 50, 51–68.
- Loudon, I. 1992. Death in Childbirth: an International Study of Maternal Care and Maternal Mortality 1800–1950. Oxford: Clarendon Press.
- Low, B. S. 1991. Reproductive life in 19th century Sweden: an evolutionary perspective on demographic phenomena. Ethology and Sociobiology, 12, 411–448.
- Lummaa, V., Haukioja, E., Lemmetynin, R. & Pikkola, M. 1998. Natural selection on human twinning. Nature, 394, 533–534. Lund, E. 1990. Pattern of childbearing and mortality in married women: a national prospective study from Norway. Journal of Epidemiology and Community Health, 44, 237–240.
- Mace, R. 1996a. Biased parental investment and reproductive success in Gabbra pastoralists. Behavioral Ecology and Sociobiology, 38, 75–81.
- Mace, R. 1996b. When to have another baby: a dynamic model of reproductive decision-making and evidence from Gabbra pastoralists. Ethology and Sociobiology, 17, 263–274.
- Mace, R. 1998. The co-evolution of human fertility and wealth inheritance. Philosophical Transactions of the Royal Society of London, Series B, 353, 389–397.
- Mace, R. In press. An adaptive model of human reproductive rate with wealth inheritance: why people have small families. In: Human Behaviour and Adaptation: an Anthropological Perspective (Ed. by L. Cronk, W. Irons & N. Chagnon). New York: Aldine de Gruyter.
- Mace, R. & Holden, C. 1999. Evolutionary ecology and crosscultural comparison: the case of matrilineality in sub-Saharan Africa. In: Comparative Primate Socioecology (Ed. by P. C. Lee), pp. 387–405. Cambridge: Cambridge University Press.
- Mace, R. & Sear, R. 1996. Maternal mortality in a Kenyan, pastoralist population. International Journal of Gynecology and Obstetrics, 74, 137–141.
- Mace, R. & Sear, R. 1997. The birth interval and the sex of children: evidence from a traditional African population. Journal of Biosocial Science, 29, 499–507.
- Moore, S. E., Cole, T., Poskitt, E., Sonko, B., Whitehead, R., McGregor, I. & Prentice, A. M. 1997. Season of birth predicts mortality in rural Gambia. Nature, 388, 434.
- Nath, D. C. & Land, K. C. 1994. Sex preferences and third birth intervals in a traditional Indian society. Journal of Biosocial Science, 26, 95–106.
- Pagel, M. D. & Harvey, P. H. 1989. Taxonomic differences in the scaling of brain weight on body weight among mammals. Science, 244, 1589–1593.
- Pennington, R. 1992. Did food increase fertility? Evaluation of !Kung and Herero history. Human Biology, 64, 497–521.
- Perls, T. T., Alpert, L. & Fretts, R. C. 1997. Middle-aged mothers live longer. Nature, 389, 133.
- Perreira, M. & Fairbanks, L. A. 1991. Juvenile Primates. New York: Oxford University Press.
- Roff, D. A. 1992. The Evolution of Life Histories: Theory and Analysis. London: Chapman & Hall. Rogers, A. R. 1993. Why menopause? Evolutionary Ecology, 7, 406–420.
- Scheper-Hughes, N. 1992. Death without Weeping: the Violence of Everyday Life in Brazil. Berkeley: University of California Press.
- Schneider, D. M. & Gough, K. 1961. Matrilineal Kinship. Berkeley: University of California Press.
- Seielstad, M. T., Minch, E. & Cavalli-Sforza, L. L. 1998. Genetic evidence for higher female migration rate in humans. Nature Genetics, 20, 278–288.
- Shanley, D. 1999. Resources, reproduction and senescence: evolutionary optimality models. Ph.D. thesis, University of Manchester.
- Simoons, F. J. 1978. The geographic hypothesis and lactose malabsorption: a weighing of the evidence. American Journal of Digestive Diseases, 23, 963–980.
- Skinner, W. 1997. Family systems and demographic processes. In: Anthropological Demography (Ed. by D. Kertzer & T. Fricke), pp. 53–95. Chicago: University of Chicago Press.
- Stearns, S. C. 1992. The Evolution of Life Histories. Oxford: Oxford University Press.
- Stearns, S. C. & Koella, J. 1986. The evolution of phenotypic plasticity in life history traits: predictions for norms of reaction for age- and size-at-maturity. Evolution, 40, 893–913.
- Strassman, B. & Dunbar, R. I. M. 1999. Human evolution and disease: putting the Stone Age in perspective. In: Evolution in Health and Disease (Ed. by S. C. Stearns), pp. 91–101. Oxford: Oxford University Press.
- Symons, D. 1979. The Evolution of Human Sexuality. Oxford: Oxford University Press. Tracer, D. 1991. Fertility related changes in maternal body composition among the Au of Papua New Guinea. American Journal of Physical Anthropology, 85, 393–406.
- Voland, E. 1988. Differential infant and child mortality in evolutionary perspective: data from late 17th to 19th century Ostfriesland (Germany). In: Human Reproductive Behaviour: a Darwinian Perspective (Ed. by L. Betzig, M. Borgerhoff-Mulder & P. Turke), pp. 253–262. Cambridge: Cambridge University Press.
- Weaver, L. T. & Beckerleg, S. 1993. Is health a sustainable state? A village study in The Gambia. Lancet, 341, 1327–1331.
- Westendorp, R. & Kirkwood, T. B. L. 1998. Human longevity at the cost of reproductive success. Nature, 396, 743–746.
- Williams, G. C. 1957. Pleiotrophy, natural selection and the evolution of senescence. Evolution, 11, 398–411.
- Wood, J. W. 1994. Dynamics of Human Reproduction: Biology, Biometry, Demography. New York: Aldine de Gruyter.
Et pour aller plus loin
Profil de nos 5000 abonnés
| Par catégorie professionnelle | |
| Médecins | 27% |
| Professions de santé | 33% |
| Sciences de la vie et de la terre | 8% |
| Sciences humaines et sociales | 12% |
| Autres sciences et techniques | 4% |
| Administration, services et tertiaires | 11% |
| Economie, commerce, industrie | 1% |
| Médias et communication | 3% |
| Art et artisanat | 1% |
| Par tranches d'âge | |
| Plus de 70 ans | 14% |
| de 50 à 70 ans | 53% |
| de 30 à 50 ans | 29% |
| moins de 30 ans | 4% |
| Par motivation | |
| Patients | 5% |
| Proche ou association de patients | 3% |
| Thèse ou études en cours | 4% |
| Intérêt professionnel | 65% |
| Simple curiosité | 23% |
Si vous n'êtes pas abonné
C'est ici
INUTILE si vous vous êtes déjà abonné, car vous le restez tant que vous ne demandez pas votre désisncription
Médecine évolutionniste (ou darwinienne)
Depuis quelques années, le problème de l'antibiorésistance, les progrès de la génomique, la redécouverte du microbiote et la prise en charge de maladies au long cours, nécessitent l'introduction d'une pensée évolutionniste dans la réflexion clinique.
Le premier diplôme universitaire intitulé "Biologie de l'évolution et médecine" a été mis en place à la faculté de Lyon en 2016.
Anthropologie / Adaptations / Sociobiologie / Culture
• Adaptation : Bel exemple d'adaptation à la plongée en apnée • Adaptation : [...]
Vous aimerez aussi ces humeurs...
Inné ou acquis ? - Les caractères individuels ont toujours alimenté de vigoureux débats sur les parts respectives [...]
Maestria des remèdes divins - Pline l’ancien dans son « Histoire naturelle » avait déjà noté les analogies entre la [...]
Soyons scientifiques ou disciplinés ou les deux - Le cancer est un sujet d'une grande gravité, c'est pourquoi il ne peut supporter la [...]
Auto-immunité - Les anti TNF font partie des avancées thérapeutiques notables de ces dernières années. [...]
Agneaux de l’agnotologie - Le danger des additifs au plomb dans l’essence a été découvert dans les [...]
Vous aimerez aussi ces humeurs...
Observance et éducation thérapeutique - L’observance thérapeutique est un sujet de grande importance pour une médecine qui se [...]
Urgences : urgence - L'urgence médicale a moins de quarante ans. Auparavant, l'exercice médical la contrôlait [...]
L'épidémiologie et le peuple - De toutes les disciplines de la médecine, l'épidémiologie est la plus complexe. Elle [...]
LUCA et le Big Bang - Depuis Galilée, qui avait tant agacé nos papes, la vulgarisation de l’astronomie a [...]
D’accord mais de mort lente - La « pression parasitaire » est l’ensemble des infections qui menacent une espèce. [...]
Vous aimerez aussi...
Danger d'extinction d'un acarien ami - Demodex folliculorum est un acarien de 0,3 mm de long qui passe sa vie profondément enfoui dans [...]
Empreinte du système immunitaire par le microbiote très tôt dans la vie - L’ empreinte du système immunitaire par le microbiote se manifeste très tôt dans la [...]
Un virus géant qui a façonné l'arbre de vie - Cette étude a identifié une famille de virus géants qui a peut-être joué un rôle clé dans [...]
Alzheimer : preuves de transmission à l'homme - Une étude sur la maladie de Creutzfeldt-Jakob a débouché sur d’étonnantes conclusions. C'est [...]
Poids de naissance et cancer - Plusieurs études montrent qu’un poids de naissance élevé est un facteur indépendant [...]
La phrase biomédicale aléatoire
Le savant complet est celui qui embrasse à la fois la théorie et la pratique expérimentale. 1° Il constate un fait ; 2° à propos de ce fait, une idée naît dans son esprit ; 3° en vue de cette idée, il raisonne, institue une expérience, en imagine et en réalise les conditions matérielles ; 4° de cette expérience résultent de nouveaux phénomènes qu'il faut observer, et ainsi de suite. L'esprit du savant se trouve en quelque sorte toujours placé entre deux observations : l'une qui sert de point de départ au raisonnement, et l'autre qui lui sert de conclusion.
― Claude Bernard
Dessins humoristiques médicaux
Les courts dialogues des manchots "givrés" du style épuré de Xavier Gorce sont [...]